Exploring the molecular mechanisms that regulate stem cell fate in complex leaves
Why Study Leaf Development?
The angiosperm leaf has a conserved function in light capture for photosynthesis, turning light energy to carbohydrates, which is the primary source for food on this planet. Not only that, but basically the waste process from the photosynthesis process in leaves results in the very air we breath. This should be reason enough to study these amazing organs, but as a developmental biologist I am interested in leaves for a very different reason, the shape of leaves can be incredible diverse while following relatively simple cellular patterns. We still have a lot to learn about how leaf shape diversity is achieved from a molecular perspective and the cellular leaf differentiation in plants. This project takes advantage of recent technologies to understand how leaf complexity is determined at a global gene expression level.
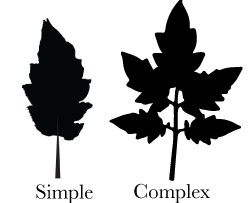
Leaf Complexity
Leaves are broadly split into two categories, simple and complex, with the complex form showing a multitude of leaflets on the leaf. Most complex leaves have a punctuated growth pattern with regions of leaflet outgrowth interrupted by regions of no outgrowth.
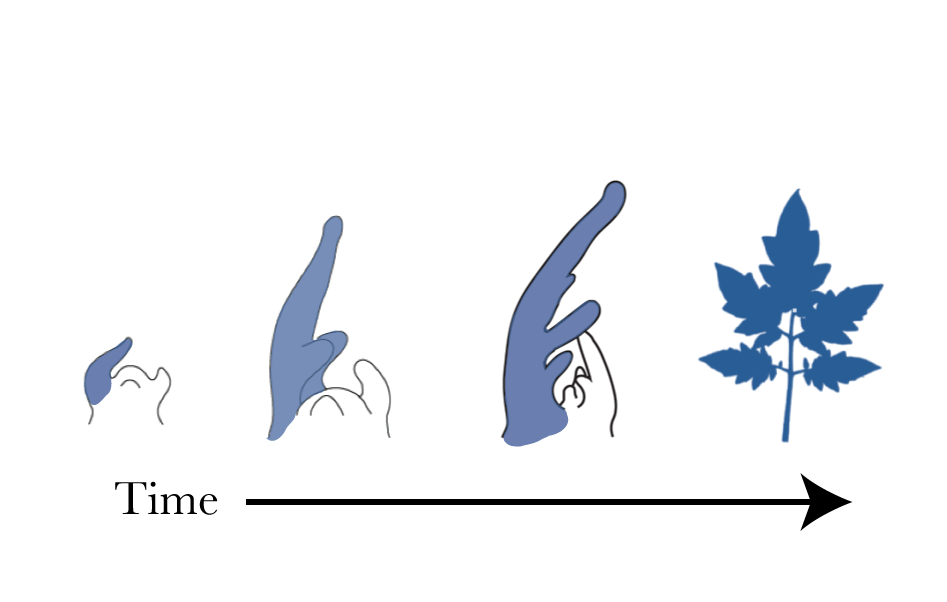
All leaves initiate from a pool of pluripotent plant stem cells called the shoot apical meristem. At these early stages of leaf development, leaves look like a uniform rod-shaped structure (see picture below), making early leaf development look very similar even across divergent plant species. At these early stages, most cells are undergoing cell division processes. In tomato leaves and many other complex leaf species, a leaflet emerges on the flanks of this rod-shaped structure. Over time, cells at the tip begin to stop dividing and instead increase in size through cell expansion creating the blade tissue. Cellular differentiation continues as cells become more specialized, for example cells differentiate plant hairs (trichomes) or stomata. Therefore during organ development the cell "life" consists of three stages 1.) Cell Division 2.) Cell expansion 3.) Cell specialization. This pattern exists in all of plant development, not just leaves, though the specific regulation of this pattern is still a hot topic in plant development.
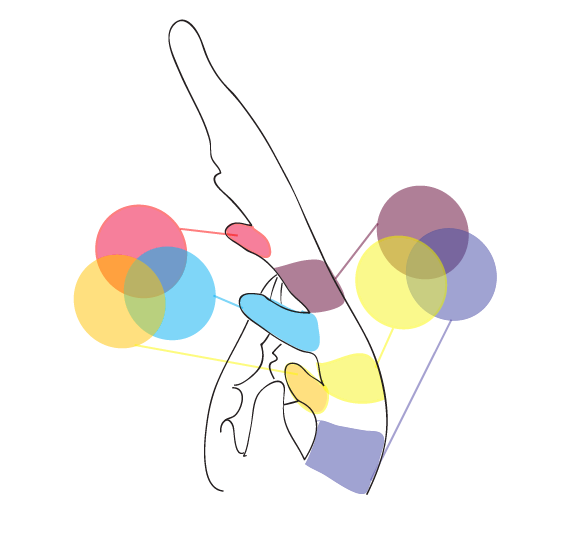
Marginal Blastozone
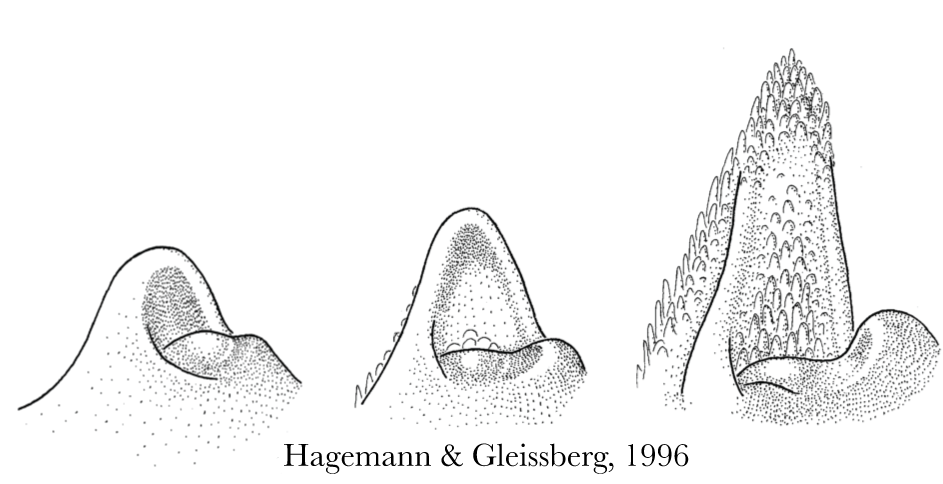
Leaflet outgrowth and blade development occurs at the Marginal Blastozone, which is the flanking sides of a developing leaf. The marginal blastozone is thought to be an area on a developing leaf that maintains meristematic or stem cell identity longer than the rest of the leaf, sometimes the marginal blastozone is even referred to as the marginal meristem. You can clearly see the marginal blastozone in the diagram below of a generic leaf developing, from Hagemann and Gleissburg, 1996. You may notice the sides of the developing leaf are uniform and smooth, lacking the specialized hair cells that are so prevalent in the last developmental stage pictured below. This is the marginal blastozone, this is where stem cell fate is maintained longest in a developing leaf, and this is where I believe the secrets to final leaf shape and cellular differentiation can be found.
Marginal Blastozone Definitions
Functional definition - Where organogenesis occurs, including leaf initiation
Histological definition - Where meristematic activity is maintained (high rates of cell division)
Mechanistic definition - Largely unknown
Therefore, in order to begin to understand more about this critical region in cell differentiation and leaf development I am looking at what genes are specifically expressed in this region.
My Project
RNA-seq
Although there is great interest in understanding how the MBR is regulated, a strategy to isolate and analyze these regions has yet to be accomplished. In recent years, sequencing technology has evolved at a rapid pace and gene expression analysis has quickly turned to RNAseq analysis to understand gene expression changes. In RNAseq analysis, RNA is isolated from specific tissue, then converted to cDNA, and Illumina sequencing of these cDNAs, allows global gene expression profiling of the specific tissue.
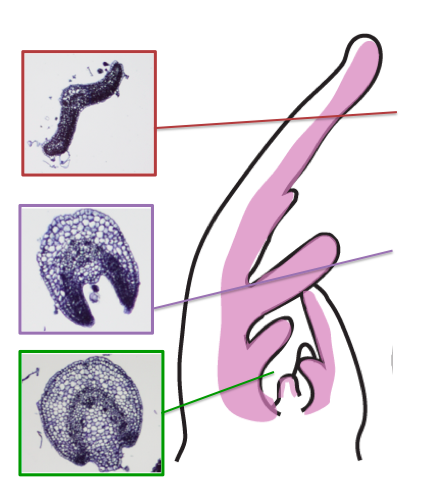
Laser Capture Micro-dissection
RNAseq analysis combined with Laser Capture Microdissection (LCM), allows the ability to isolate and analyze the gene expression profile of tissue regions previously unattainable, like the MBR in small early developing leaves. Solanum lycopersicum (tomato), with its large accessible meristem and young leaves, a recent sequenced genome, and a tomato mutant that has early MBR differentiation, trifoliate, make tomato the perfect system for this project. The combination of these approaches allow the exploration of how cells in these MBR regions maintain competency and differentiate t o direct the vast quantity of leaf shape diversity achieved in the plant kingdom.